Социальные амебы Dictyostelium discoideum делятся на три «пола», каждый из которых может спариваться с любым из двух других. Оказалось, что половая принадлежность амеб определяется единственным генетическим локусом, содержащим 1, 2 или 3 гена. Ключевую роль играют гены двух типов, непохожие ни на какие ранее известные гены. Для половой совместимости необходимо, чтобы один из партнеров имел ген первого типа, другой — второго.
Амебы Dictyostelium в последнее время стали популярным лабораторным объектом. Об их способности образовывать многоклеточные плодовые тела из множества индивидуальных организмов, многие из которых жертвуют жизнью ради «общего блага», рассказано в заметке Амёбы-мутанты не позволяют себя обманывать («Элементы», 06.10.2009).
Удивительные особенности диктиостелиума не исчерпываются сложным поведением при образовании плодовых тел. Половое размножение у этих амеб тоже протекает в высшей степени необычно. У диктиостелиума не два, а три «пола», или типа спаривания (mating types). Само по себе это еще не очень удивительно: подобная «многополость» известна у некоторых низших эукариот, в том числе у грибов и инфузорий. Если половые клетки не различаются по размеру и строению (см. Isogamy), то есть не делятся на крупные яйцеклетки и мелкие спермии, то число «полов» не обязано быть равным двум. Однако у диктиостелиума половое размножение обставлено дополнительными причудливыми «ритуалами», включающими сложное социальное поведение и каннибализм.
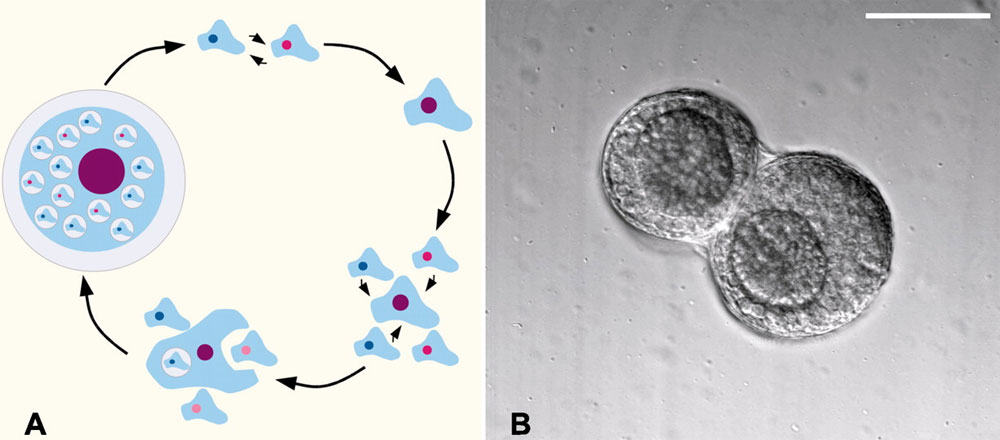
В благоприятных условиях гаплоидные одиночные амебы размножаются делением. Столкнувшись с нехваткой пищи, они могут перейти к половому размножению. Для этого должны встретиться две амебы, относящиеся к разным «полам». Каждый из трех полов (I, II и III) может скрещиваться с любым из двух других. Две гаплоидные амебы сливаются и образуют крупную диплоидную амебу — зиготу. После этого начинается самое интересное. Зигота выделяет сигнальное вещество — циклический аденозинмонофосфат (цАМФ), привлекающий гаплоидных амеб. Это же вещество используется амебами в качестве сигнала «ползите все сюда» при образовании скоплений, из которых потом формируется плодовое тело.
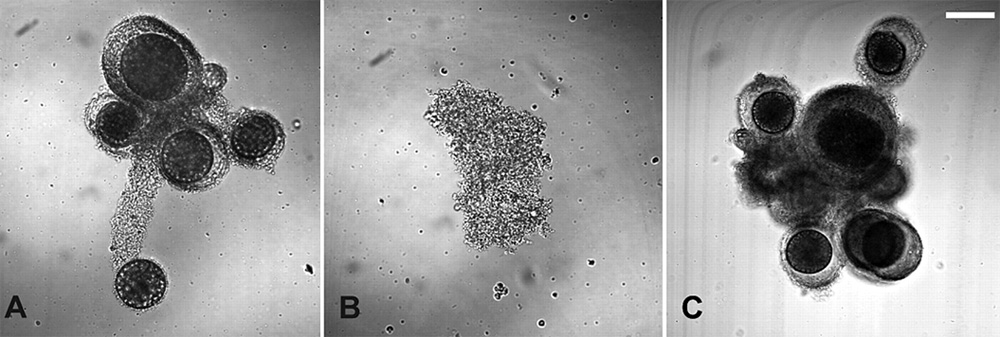
При образовании плодового тела 80% амеб превращаются в споры, получая шанс передать свои гены следующим поколениям, а 20% жертвуют собой: их тела идут на построение ножки плодового тела. Совсем другой расклад получается, когда амебы доверчиво подползают к зиготе. Подманив, словно сирена, множество гаплоидных амеб, зигота заглатывает их путем фагоцитоза и переваривает. При этом ее размер, естественно, увеличивается. В итоге получается гигантская клетка — макроциста, которая может быть в 500–1000 раз крупнее одиночной амебы. До того как быть съеденными, одиночные амебы, окружившие зиготу, строят вокруг будущей макроцисты прочную трехслойную стенку из целлюлозы. Таким образом, зигота использует маленьких гаплоидных амеб не только как пищу, но и как рабочую силу.
При наступлении благоприятных условий макроциста «прорастает», и из нее выходят сотни маленьких гаплоидных амеб. Все они, конечно, являются потомками зиготы, а не тех несчастных, которые были ею съедены. По-видимому, перед выходом потомства зигота сначала претерпевает мейоз, а затем множество последовательных митозов (хоть это и не доказано окончательно).
Предполагается, что механизм формирования макроцист эволюционно древнее механизма формирования плодовых тел, причем второй, возможно, произошел от первого.
Несмотря на то что многие лаборатории уже вовсю используют диктиостелиума в качестве модельного объекта для изучения социального поведения и химической коммуникации, многие аспекты жизни этого организма остаются загадочными. Например, до сих пор не было известно, от чего зависит пол амеб, какие гены определяют принадлежность амебы к одному из трех типов спаривания. Британские и японские ученые сообщили о разгадке этой тайны в последнем выпуске журнала Science.
Авторы целенаправленно искали в геноме диктиостелиума гены, имеющиеся у одних полов и отсутствующие у других. Геном пола I прочтен, что позволило изготовить ДНК-микрочип с образцами последовательностей 8500 генов из примерно 10 500, обнаруженных в геноме пола I. При помощи этого микрочипа были исследованы геномы 10 диких штаммов диктиостелиума, относящихся к полам I и II. В итоге был выявлен один-единственный ген в пятой хромосоме, который есть у всех амеб пола I и отсутствует у всех амеб пола II. Авторы назвали этот ген matA. Он кодирует короткий (длиной в 107 аминокислот) белок, непохожий ни на какие известные белки.
Чтобы убедиться в том, что обнаруженный белок действительно определяет половую принадлежность амеб первого пола, авторы удалили этот ген из их генома. В результате амебы полностью утратили способность спариваться и образовывать макроцисты с любыми амебами независимо от их пола. Когда ген вернули на место, способность спариваться с амебами второго и третьего полов восстановилась.
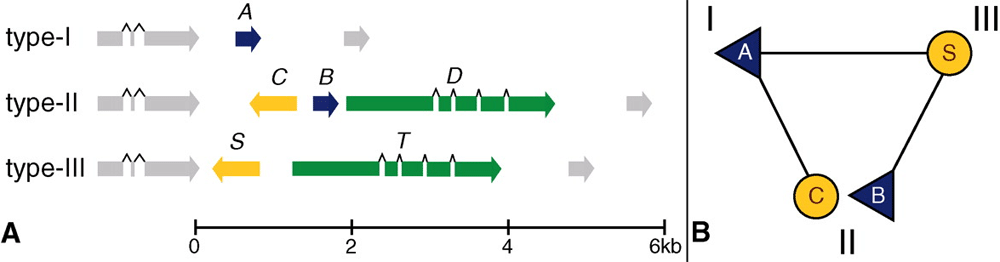
В геноме пола I по обе стороны от matA находятся гены, имеющиеся у всех трех полов и занимающие одинаковые позиции в хромосоме. Это обстоятельство позволило исследовать соответствующий участок пятой хромосомы у всех трех полов при помощи метода ПЦР (см. Полимеразная цепная реакция). Оказалось, что у пола II между этими общими для всех полов генами находится не один (как у пола I), а три гена, которые получили названия matB, matC и matD. Первый из них гомологичен гену matA, однако аминокислотные последовательности белков, кодируемых генами matA и matB, совпадают лишь на 60%. Ген matC не похож на другие известные гены, ген matD отдаленно напоминает одно из известных семейств генов, участвующих в слиянии гамет.
При помощи генно-инженерного эксперимента удалось показать, что гены matB, matC и matDдействительно определяют половую принадлежность амеб второго пола. Авторы удалили у амеб первого пола ген matA, а затем вставили в их геном эти три гена. Получившиеся мутанты вели себя как амебы второго пола: они спаривались с полами I и III и не могли образовать макроцисты с полом II.
Аналогичным образом были выявлены гены, определяющие половую принадлежность амеб третьего пола. Таких генов оказалось два: matS и matT, причем первый из них сходен с matC, а второй — с matD.Ничего похожего на matA и matB в геноме третьего пола не обнаружилось.
Таким образом, локус типа спаривания (mating-type locus) у первого и третьего полов не содержит сходных элементов, а у второго пола он похож на комбинацию двух других.
Дальнейшие эксперименты показали, что три гена, находящиеся в локусе типа спаривания у амеб второго пола, выполняют разные функции. Один из них, matB, позволяет спариваться с третьим полом; другой,matC, — с первым. Ген matD не влияет на половую принадлежность, однако его наличие при некоторых скрещиваниях увеличивает число образовавшихся макроцист. Возможно, matD повышает вероятность слияния гаплоидных амеб и образования зигот.
Из двух генов, находящихся в локусе типа спаривания у амеб третьего пола, ключевым оказался ген matS. Именно от него зависит способность к спариванию с двумя другими полами. При спаривании с амебами второго пола решающую роль играет взаимодействие между генами matS и matB. Ген matT не участвует в определении пола; его функции остались неизвестны.
Таким образом, в системе определения пола у диктиостелиума можно проследить определенную логику. У полов I и III половая принадлежность определяется единственным геном — соответственно, matA иmatS. Для совместимости необходимо, чтобы один из партнеров имел ген matA или его гомолог, а другой — ген matS или его гомолог. Амебы второго пола имеют сразу два «половых гена» matB и matC, являющиеся гомологами matA и matS. Наличие гомолога matA позволяет второму полу скрещиваться с третьим, гомолога matS — со вторым полом. Почему амебы второго пола не могут при этом скрещиваться друг с другом, пока не ясно.
Авторы предполагают, что у предков диктиостелиума было только два пола (I и III), а пол II появился позже в результате объединения регуляторных генов двух исходных полов.
Расшифровка механизма определения пола у диктиостелиума должна существенно облегчить разнообразные генетические эксперименты с этим интересным лабораторным объектом.
Источник: Gareth Bloomfield, Jason Skelton, Alasdair Ivens, Yoshimasa Tanaka, Robert R. Kay. Sex Determination in the Social Amoeba Dictyostelium discoideum // Science. 2010. V. 330. P. 1533–1536.
См. также:
Амёбы-мутанты не позволяют себя обманывать, «Элементы», 06.10.2009.
Александр Марков